J'ai pris le parti de n'introduire que 3 "termes techniques" : "chromosome", "gène" et "allèle", qui sont expliqués en introduction.
Pour deux raisons :
- ça permet une compréhension logique et ça évite les confusions (notamment dans le cas des dilutions et des masques jaunes)
- ça permet de comprendre tous les cas de figure de croisements, y compris les plus subtils (du genre : "Comment puis-je obtenir des "ailes en dentelle" avec des cinnamon et des ino?")
- ça correspond à la biologie qui se cache derrière la génétique.
Les autres termes qui sont nécessaires pour comprendre les liens entre les différentes couleurs ("recessif" et "dominant") sont expliqués au fur et à mesure des exemples.
Pour ceux qui ont envie d'apprendre d'autres termes de génétique, il y a un petit lexique en bas. Mais ces termes ne sont pas nécessaires pour comprendre les principes et prédire les résultats des croisements.
Ceux qui ont l'habitude de parler de mutations et de facteurs de couleur s'y retrouveront sans problème car le principe est le même, en plus simple. Pour les débutants, une fois que vous aurez compris le principe de base, vous aurez juste l'impression de faire des mélanges de peinture sur une palette en plumes. C'est très amusant!
Et j'espère que certains pourront aussi trouver cela utile...
Les perruches ondulées vivent à l'état sauvage en Australie. Les oiseaux sauvages sont très semblables entre eux, et servent de référence. Voici une photo (oiseau du fond):

Les mâles et les femelles ont un plumage de même couleur à l'état sauvage, mais chez les mâles, la cire du bec (qui entoure les narines), est bleue intense. Chez les femelles, elle est brune en période de reproduction, et blanc bleuté en dehors.
Les jeunes ont un plumage un peu plus terne que les adultes, avec des ondulations noires qui couvrent tout le front. Leur cire est rose-violet chez les mâles, et blanc bleuté chez les femelles. Leurs yeux sont noirs, car l'iris est coloré de noir.
Deux pigments sont responsables de la couleur de cet oiseau : la psittacine qui est jaune, et la mélanine qui est noire. Le vert est un mélange du jaune de la psittacine et d'un bleu qui n'est pas dû à un pigment. Il est obtenu par diffraction de la lumière sur les plumes contenant de la mélanine.
Mouillé, le ventre de cet oiseau sera marron : jaune (psittacine) plus noir (mélanine). Si il n'y a pas de jaune, un ventre bleu mouillé apparaît gris. On le voit bien sur cette photo prise après un bain:

En captivité ont été sélectionné différentes couleurs (comme l'oiseau de premier plan sur la photo 1).
La couleur d'une perruche est déterminée par sa capacité à fabriquer :
- des pigments, psittacine jaune et mélanine noire
- des plumes capables de diffracter la lumière.
Cette capacité dépend directement de ses gènes. Chez certains oiseaux, comme les canaris, l'environnement peut aussi jouer (nourris de caroténoïdes, il deviennent plus jaunes ou rouges), mais ce n'est pas le cas des psittacidés.
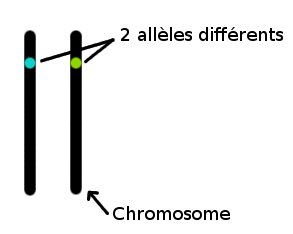
Les gènes sont des portions de "chromosomes", une longue molécule d'ADN. L'oiseau reçoit toujours une moitié de ses chromosomes de son père, et une moitié de sa mère, selon le schéma ci-dessous.

Chaque oiseau a donc deux exemplaires de chaque chromosome, et ainsi de chaque gène. On appelle ces "exemplaires" des allèles. Ils peuvent être identiques ou différents. Quand une cellule se divise, par exemple pour la croissance, elle doit recopier ses chromosomes, de sorte que chaque cellule-fille ait toute l'information génétique. Parfois se produit une erreur au cours de la copie, c'est une mutation. Si cette mutation a lieu dans les cellules reproductrices, alors elle peut être transmise à la descendance. Un nouvel allèle est apparu!
Chaque gène peut avoir plusieurs allèles dans la population, même si un oiseau ne peut en porter que deux.
Dans tous les allèles que l'on connaît, certains sont apparus par mutation il y a potentiellement très longtemps, mais sont restés rares ou très rares dans les populations sauvages. D'autres au contraire sont apparus récemment, au cours de la domestication. On peut rarement faire la différence entre les deux.
Quoiqu'il en soit, la consanguinité (=le croisement d'individu apparentés, possédant le plus d'allèles en commun, hérités de leurs parents proches communs), permet de révéler les allèles rares.
Dans la nature, la grande majorité des oiseaux possède les mêmes allèles de couleur. Ils sont presque tous semblables. Mais parfois, par hasard, nait un individu qui possède une caractéristique différente des autres, comme un corps bleu au lieu de vert (pas de jaune). Cela cela permet d'identifier un gène, ici impliqué dans la fabrication du pigment jaune. On donne alors à ce gène le nom de la couleur qui vient d'apparaître.
Dans notre cas ce sera "bleu".
Il faut distinguer néanmoins les deux allèles : celui qui permet de fabriquer du jaune, le plus fréquent dans la population sauvage et celui qui ne le permet pas. Le second se nommera "b" pour bleu et le premier b+ par convention (+ signifie "type sauvage").
Pour les noms des allèles, nous utiliserons les noms donnés sur la page wikipedia consacrée aux couleurs de perruches (voir sources), faciles à retrouver et très proches des standards internationnaux (si ils ne sont pas directement les standards internationnaux). Pour les noms des gènes eux-même et des couleurs, nous utiliserons des traductions en français, c'est plus facile.
BLEU (on commence par deux allèles, b et b+, on reprendra plus tard avec la totalité des allèles connus) - influe la capacité à synthétiser du pigment jaune
L'oiseau ayant deux allèles de chaque gène, il y a donc trois cas de figure : b/b, b+/b et b+/b+ Ces oiseaux auront des couleurs différentes :-b/b est bleu
-b+/b est vert
-b+/b+ est vert
Il suffit donc d'un seul allèle "b+" pour que l'oiseau puisse fabriquer de la psittacine et soit vert. Le vert est donc "dominant" sur le bleu, lequel est "récessif" par rapport au vert. Par convention on note les allèles correspondant aux caractères dominants en majuscule. C'est facile quand il n'y en a que deux, sinon on laisse tout le monde en minuscules.
Si l'on a un oiseau vert, il est donc impossible de savoir si il est B+/b ou B+/B+ à moins qu'il soit lui-même issu d'au moins un parent bleu.
Un oiseau qui porte un allèle pour un caractère récessif, mais sans l'exprimer par ce qu'il n'en a qu'un seul exemplaire, est dit "porteur" du caractère récessif.
Un B+/b est donc "porteur de bleu".
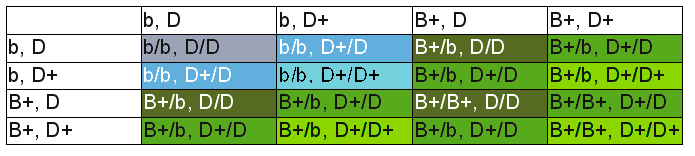
Dans le cas d'un croisement entre un porteur de bleu et un bleu, on peut voir sur le tableau suivant (à gauche) deux possiblités (en tête de ligne et colonne on a les allèles portés par les deux parents, les couleurs des cases représentent la couleur du corps des descendants en fonction de leurs allèles):

De tels tableaux de croisements permettent de construire des hypothèses et de faire des prévisions des résultats des croisements. Avec deux oiseaux verts, on peut imaginer les cas où les deux sont B+/B+, où les deux sont B+/b et où l'un est B+/B+ et l'autre B+/b
FONCÉ (deux allèles D+ et D) - influe la structure des plumes et la façon dont elles renvoient la lumière
Intéressons-nous maintenant à une autre variation de couleur : foncé.On la trouve dans la série verte ou la série bleue. Il y a deux allèles, D et D+ (celui possédé le plus fréquemment dans la population sauvage). Ils possèdent une particularité intéressante:
un oiseau D+/D+ est clair "sauvage", un D+/D est foncé et un D/D très foncé (au moins chez les verts). On voit donc que quand l'oiseau porte deux allèles différents, il a une couleur intérmédiaire entre les deux extrèmes. De tels allèles sont dits co-dominants.
Si l'on combine le gène "Bleu" avec le gène "foncé", on obtient toute une gamme de couleurs :
- b+/b, D+/D+ ou b+/b+, D+/D+ : vert clair "type sauvage"
- b+/b, D+/D ou b+/b+, D+/D : vert foncé
- b+/b, D/D ou b+/b+, D/D : olive
- b/b, D+/D+ ou b+/b+, D+/D+ : bleu clair "type sauvage"
- b/b, D+/D : cobalt
- b/b, D/D : mauve (un gris violacé)

Oiseau Cobalt

Oiseau Olive
Voici un exemple de croisement de deux parents qui ont les deux allèles différents pour les deux gènes (les allèles des parents sont en tête de ligne et de colonne, les deux gènes séparés par une virgule, le tout dans toutes les combinaisons possibles - la couleur de la case représente en gros la couleur du corps de chaque type de descendant):
Pour plus de facilité, il vaut mieux faire des tableaux simples, à 2 lignes et 2 colonnes. On fait ainsi un seul tableau par gène, ce qui permet de traiter autant de gènes que l'on veut!
Néanmoins, il y a une raison particlulière à traiter les gènes "bleu" et "foncé" sur le même tableau : ils sont physiologiquement placés sur le même chromosome. Ce qui signifie que la proportion de petits de chaque couleur ne dépendra pas seulement des allèles portés par les parents, mais aussi de leur combinaison sur les différents chromosomes.
Si on jette un oeil au schéma ci-dessous, qui représente deux chromosomes, chacun portant un allèle du gène bleu, et un allèle du gène foncé :
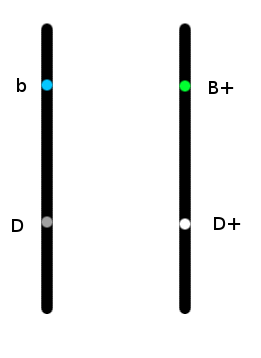
On se rend compte que les allèles sont liés physiquement. Cet oiseau aura une très grande probabilité de transmettre ensemble les allèles D+ et B+, et une très faible probabilité (moins de14% d'après wikipedia) de transmettre ensemble les allèles D et B+. Il faut pour cela que les chromosomes s'échangent des morceaux au moment de la fabrication des gamètes (ovules et spermatozoïdes).
Donc cette considération peut être à prendre en compte pour augmenter ses chances de produire des oiseaux avec des combinaisons spéciales entre les gènes "bleu" et "foncé". Pour faciliter le repérage, quand on sait grâce aux croisements sur quel chromosme sont placés les allèles, on peut noter les allèles possédés par les oiseaux avec les deux allèles de chaque côté de la barre oblique.
Par exemple, un vert clair "type sauvage" sera noté : b+_D+/b_D+ ou b+_D+/b+_D+.
Un vert foncé b+_D+/b_D ou b+_D/b_D+ ont les mêmes allèles mais n'auront pas la même probablité d'avoir des descendants de chaque couleur.
Il faut faire des tableaux de croisement pour se rendre compte des changements.
OPALINE (deux allèles, Op+ et op) - influe la distribution du pigment noir sur le plumage
Voilà un gène particulier, du fait qu'il se trouve sur l'un des chromosomes sexuels. Chez l'homme, les chromosomes sexuels sont "XX" chez la femme (toutjours deux chromosomes, un hérité du père, un de la mère), "XY" chez l'homme. Y est un tout petit chromosome qui détermine la masculinité. Chez les oiseaux, les mâles sont ZZ et les femelles WZ (on ne dit pas X et Y pour ne pas confondre avec le système des mammifères).Opaline est un gène porté par le chromosome Z. Les femelles n'en ont donc, exceptionnellement, qu'un seul et unique exemplaire.
Elles sont Zop/W (le dessin noir sur leurs dos et leur ailes est modifié comme sur la photo ci-dessous), ou bien ZOp+/W (Op+ renvoie au type sauvage : pas de modification du dessin).

Chez les mâles, on a un schéma "classique" : Zop/Zop provoque le dessin "opaline" ZOp+/Zop laisse un dessin "type sauvage", donc la couleur "opaline" est récessive par rapport au dessin "type sauvage".
BLEU (tous les allèles : b+, b1, b2, byf et bgf) - influe la synthèse du pigment jaune
Ce traître de gène bleu, qui paraissait si simple tout à l'heure, est en réalité un peu plus complexe qu'il en a l'air. En effet, il ne se contente pas d'"éteindre" la synthèse de jaune, il peut simplement l'empêcher de se produire sur une partie du corps de l'oiseau.On lui connaît 5 allèles, dont voici les effets quand ils sont présents en double exemplaires :
- b+, le type sauvage, qui permet la synthèse de pigment jaune sur tout le corps
- b1, supprime le jaune de tout le corps
- b2, supprime aussi le jaune de tout le corps
- byf, supprime le jaune du bas du ventre
- bgf, supprime le jaune partout sauf sur la tête.
Les différentes combinaisons de ces allèles donnent des couleurs très variées :
- b+/b1 : couleur verte "type sauvage"
- b+/b2 : couleur verte "type sauvage"
- b+/byf : couleur verte "type sauvage"
- b+/bgf : couleur verte "type sauvage"
Le vert est dominant sur les autres couleurs. Les oiseaux dits "de série verte" possèdent donc au moins un allèle b+. Tous ceux qui n'en ont pas sont dits "de série bleue" (même si ils peuvent aussi avoir du vert, comme on va le voir plus bas...)
- b1/b1 : pas de jaune du tout, oiseau bleu.
- b2/b2 : pas de jaune du tout, oiseau bleu
- b1/b2 : "masque jaune type 1", le pigment jaune n'est présent pratiquement que sur la tête.
- b1/byf : "masque jaune type 2 intense" : le jaune est dilué sur le corps, le ventre est turquoise tirant vers le vert, et la tête est jaune un peu clair.
- b2/byf : "masque jaune type 2 léger" : le jaune est dilué sur le corps, le ventre est turquoise tirant vers le bleu, et la tête est jaune un peu clair.
- byf/byf : "masque jaune type 2 moyen" : le jaune est dilué sur le corps, le ventre est turquoise moyen, et la tête est jaune un peu clair.
- b1/bgf : "masque doré intense" : le jaune est un très peu dilué sur le bas du ventre, qui tire un peu vers le bleuté, le reste du corps est vert.
- b2/bgf :"masque doré léger" : le jaune est dilué sur le bas du ventre, qui tire vers le bleu.
- bgf/bgf: "masque doré moyen" : le jaune est un peu dilué sur le bas du ventre, qui tire vers le bleu, mais moins que chez le b2/bgf.
Les allèles b1 et b2 sont dits "superdominants". C'est un cas qui ne se rencontre pas très fréquemment, où les deux allèles se complètent pour restaurer une partie de la fonction du gène (ici, fabriquer du jaune), alors que tous seuls ils ne font rien ou pas grand chose. On ne peut pas différentier b1 et b2, mais on peut savoir qu'ils sont là quand un masque jaune type 1 appara&icric;t.
Accoupler deux "masques jaunes de type1" ne peut donc donner que les résultas suivants (les cases correspondant aux parents sont en têtes de lignes et de colonnes, celles correspondants aux descendants sont jaunes quand il y a un masque jaune, blancs quand le masque reste blanc) :

Voici une image d'un oiseau masque jaune type 1:

De la même façon, bgf et b1 sont superdominants, c'est à dire que si l'oiseau possède ces deux allèles ensemble (bgf/b1), il a davantage de jaune que si il ne possède que deux exemplaires du même allèle (bgf/bgf ou b1/b1). Par contre, Bgf est partiellement dominant sur b2, puisque quand les deux allèles sont présents ensemble, le masque doré est "léger".
Et enfin, Byf et b1 sont superdominants également. Quand l'oiseau possédant les deux allèles ensemble (byf/b1) il exprime davantage de jaune que ceux qui ne possèdent que deux exemplaires du même allèle. Byf est quant à lui partiellement dominant sur b2.
C'est plus clair avec un schéma (où l'on reprendra les nomenclatures "simple facteur" et "double facteur", pour montrer que le même nom peut désigner deux oiseaux bien différents):
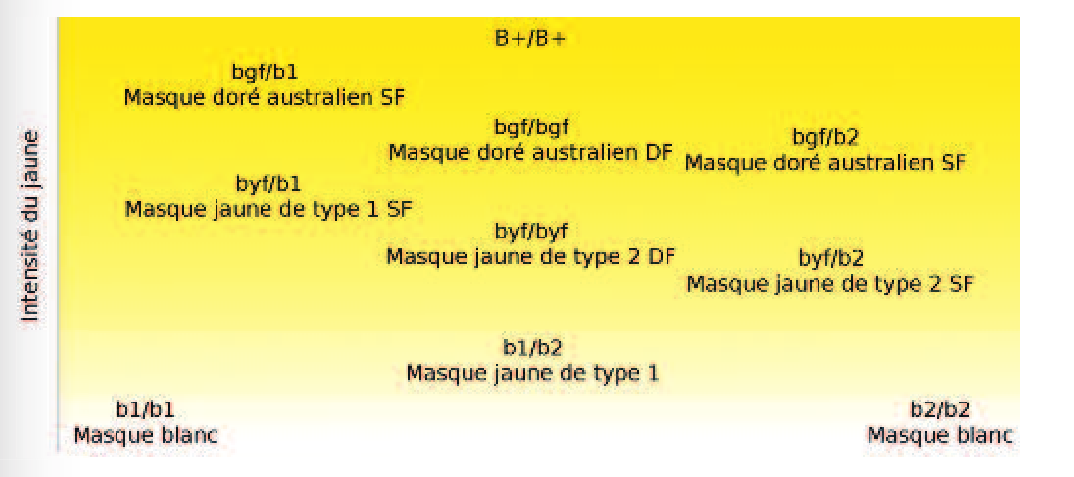
Photo de masque jaune type 2 "intense" (le flash la fait paraître un tout petit peu plus claire qu'elle en l'est en réalité)

On ne peut pas utiliser la convention de la majuscule pour la dominance, parce qu'avec ces quatre-là, ce serait trop compliqué. Il est également important de noter que le jaune a tendance à s'intensifier après la première mue. On ne peut donc vraiment voir les couleurs telles qu'elles sont décrites que lorsque l'oiseau est jeune, après, les différentier devient de plus en plus difficile. Dernier point : les spécialistes tels que Peter Bergman et Otto Völker, après avoir observé des plumages de perruches ondulées sous lumière ultraviolette (que les oiseaux perçoivent), ont remarqué que seuls certaines parties des plumes colorées en jaune renvoyaient les UV. Il y a donc probablement non pas un mais deux pigments jaunes chez les ondulées. Le pigment jaune qui renvoie aussi la lumière UV dessine des motifs semblables à ceux observés en jaune chez les masques jaunes de type 2 et masques dorés australiens. Ce pigment serait laissé plus ou moins intact par les allèles byf et bgf, qui supprimeraient plutôt le pigment non réfléchi par les UV.
DILUTION (quatre allèles, dil+, dilgw, dilcw, et dild)- influe la synthèse du pigment noir (quantité)
Maintenant que l'on a acquit tout le vocabulaire nécessaire à la compréhension de la génétique, interressons-nous au gène "dilué", "dil" pour les intimes. Il détermine la dilution de la mélanine, le pigment noir. Non dilué, l'oiseau a des marques noires sur les ailes ("ondulations"). Avec un des trois autres allèles, ce noir est dilué en gris plus ou moins intense.Donc en tout il existe 4 quatre allèles, nommé en fonction de la couleur de l'oiseau quand il possède deux allèles identiques:- dil+ : forme "sauvage"
- dilgw : ailes grises (grey wing)
- dilcw : ailes claires (clear wings)
- dild : dilué
Nous allons maintenant regarder toutes les combinaisons possibles, et voir la couleur qu'elles donnent :
dil+/dil+ : type sauvage, ondulations noires.
dil+/dild : type sauvage
dil+/dilcw : type sauvage
dil+/dilgw : type sauvage
L'allèles dil+ est donc dominant sur les trois autres.
dilgw/dilgw : ailes grises
dilgw/dilcw : fullbody : intermédiaire entre ailes grises et ailes claires : les deux allèles sont codominants
dilcgw/dild : ailes grises
L'allèle dilgw est donc récessif par rapport à dil+, codominant par rapport à dilcw et dominant par rapport à dild.
dilcw/dilcw : ailes claires
dilcw/dild : ailes claires
L'allèle dilcw est donc dominant par rapport à dild.
dild/dild : dilué (encore plus clair que "ailes claires, couleur du corps plus claire aussi)
INO (deux allèles ino+, inocl et ino) - influe la synthèse du pigment noir
Ce gène est lui aussi situé sur le chromosome sexuel Z. Il existe trois allèles : le "sauvage" ino+, qui est le plus fréquent dans la population sauvage. Il permet la synthèse de la mélanine sur tout le corps de l'oiseau, donc un ventre coloré (bleu ou vert selon le cas), et des ailes portant des ondulations noires. Les autres allèles sont:- inocl : quand un mâle en possède deux exemplaires (pour les femelles, elles ne peuvent pas en posséder plus d'un puisque nous sommes sur un chromosome sexuel), il est de couleur "texas clearbody", avec de la mélanine sur les ailes uniquement.
- ino : quand un mâle en possède deux exemplaires (ou les femelles un), il est de couleur "Ino", sans aucune trace de mélanine (attention à la confusion entre le nom de l'allèle et le nom de la couleur... on va mettre une majuscule au nom de la couleur pour ne pas les confondre).
Jettons un oeil sur l'ensemble des possibilités. D'abord les femelles.
- Zino+/W : "mélanine sur le corps et les ailes type sauvage"
- Zinocl/W : Texas clear body
- Zino/W : Ino
Les mâles :
- Zino+/Zino+ : "mélanine sur le corps et les ailes type sauvage"
- Zino+/Zinocl : "mélanine sur le corps et les ailes type sauvage"
- Zino+/Zino : "mélanine sur le corps et les ailes type sauvage"
Il suffit donc d'un seul allèle ino+ pour fabriquer de la mélanine : la forme sauvage est dominante sur les formes ino et texas clear body.
- Zinocl/Zinocl : Texas clear body
- Zinocl/Zino : Texas clear body
- Zino/Zino : Ino
Texas clear body est donc dominant sur Ino.

Deux Texas clearbody (série verte et série bleue). Photo copyright : Didier Mervilde


Un lutino. (Photo copyright : Pascale Maitre)
De quelle couleur seront exactement les perruches "Ino"? ça dépend de leur capacité à produire du pigment jaune, et donc de leurs allèles au niveau du gène "bleu". Si elles le peuvent, elles seront plus ou moins jaunes (Lutino), sinon elles seront blanches (Albino). Leurs yeux seront toujours rouges.
Les Texas clear body seront de la même façon blanches avec des ondulations noires sur les ailes, ou bien jaunes avec des ondulations noires sur les ailes.
Par contre, quels que soient leurs allèles au niveau des autres gènes responsables de la fabrication de la mélanine, les oiseaux ne fabriqueront aucune mélanine.
Il y a néanmoins une exception, c'est quand un certain allèle du gènes "Cinnamon" vient s'en mêler. (on va en parler après avoir regardé "Cinnamon" de plus près...)
CINNAMON (deux allèles cin+ et cin) - influe la synthèse du pigment noir
Cinnamon est un gène très spécial. Il transforme la mélanine noire en mélanine plus marron et plus claire. Il est situé sur le chromosome sexuel Z, comme ino et opaline. Mais en plus, sur ce chromosome, il est très proche physiquement de ino, ce qui peut avoir une importance quand on cherche à combiner ces deux mutations pour obtenir le très rare et très joli "Ailes en dentelles".Comme pour les gène opaline et ino[/i], il y a trois possibilités pour les mâles, et deux pour les femelles. Les femelles ne peuvent être que Zcin+/W (type sauvage) ou Zcin/W (Cinnamon)
Les mâles seront soit :
- Zcin+/Zcin+ : "type sauvage"
- Zcin+/Zcin : "type sauvage"
- Zcin/Zcin : Cinnamon

Là où ça se complique un petit peu, c'est quand on s'aperçoit que les femelles Zino_cin/W, Zcin/W, et les mâles Zino_cin/Zino_cin, au lieu d'être "Ino", c'est à dire incapables de fabriquer de la mélanine, possèdent en fait de très fins lisérés beiges sur les ailes. L'allèle cin restaure en partie l'incapacité des oiseaux "Ino" à fabriquer de la mélanine!

(Photo copyright : Didier Mervilde)

Comme les gènes cinnamon et ino sont très proches physiquement sur leur chromosome, il n'est toutefois pas évident de combiner les allèles rares ino et cin. Il a fallut réaliser plusieurs croisement en utilisant des mâles Zcin+_ino/Zcin_ino+, pour y arriver, grâce à un phénomène qui se passe lors de la fabrication des spermatozoïdes (ou des ovules, mais pas sur les chromosomes sexuels des femelles parce que W est trop petit), où les chromosomes s'échangent des morceaux. Avec un schéma, ce sera plus clair :
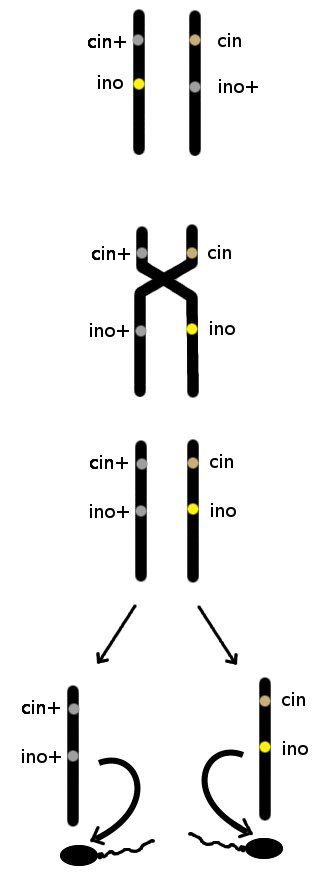
Donc pour avoir des "ailes en dentelles", il faut soit croiser des individus qui ont déjà les allèles correctement ordonnés, soit utiliser des mâles porteurs ino et cinnamon (Zcin+_ino/Zcin_ino+) puis de croiser leurs descendants ente eux et avoir beaucoup de patience (l'échange de morceaux de chromosomes chez les males porteurs intervient dans environ 36% des cas).
Par contre, des ailes en dentelles croisées ensemble donneront toujours des ailes en dentelles puisqu'elles ne portent plus d'exemplaire de type "sauvage" ni pour le gène ino, ni pour le gène cinnamon.
ARDOISÉ (deux allèles, sl+ et sl) - influe sur la structure des plumes.
Ardoisé, ou slate en anglais (d'où le nom du gène), est un gène qui modifie la structure des plumes et la façon dont elles diffractent la lumière. C'est le dernier gène connu porté par le chromosome Z, et donc lié au sexe. Il y a donc deux possiblités pour la femelle:Zsl+/W : "type sauvage", le bleu est diffracté normalement
Zsl/W : le bleu diffracté est plus sombre et moins vif, le rendant vert-gris sombre (jaune+bleu-gris sombre) ou bleu-gris sombre. La couleur réelle de l'oiseau dépend de la présence d'allèles modifiant la couleur du plumage au niveau d'autres gènes, comme "bleu" ou "foncé".
Pour les mâles, trois possiblités :
Zsl+/Zsl+ : "type sauvage", le bleu est diffracté normalement
Zsl+/Zsl : "type sauvage", le bleu est diffracté normalement
Zsl/Zsl : le bleu diffracté est plus sombre et moins vif. La couleur réelle de l'oiseau dépend de la présence d'allèles modifiant la couleur du plumage au niveau d'autres gènes, comme "bleu" ou "foncé".
Il semble difficile de produire des opalines ardoisées, car les deux gènes sont assez proches sur les chromosomes. Il faut donc attendre les échanges aléatoires de morceaux entre chromosomes (voir Ino et cinnamon.
VIOLET (deux allèles, V+ et V) - influe sur la structure des plumes
Violet est un gène porté par un chromosome non sexuel. Comme "Ardoisé", il modifie la structure des plumes.Il y a donc trois possiblités pour les deux sexes :
- V+/V+ : "type sauvage"
- V+/V : sur du vert, il fonce un peu la tonalité. Sur du bleu, l'effet varie en fonction de la présence des divers allèles de "foncé".
- V/V : Les verts sont en principe encore un peu plus foncés que les V+/V, et les bleus varient aussi, de la même façon.
Voici un tableau des différentes variations (autant que possible, les codes pantome ont été respectés):


Individu violet
L'oiseau de la photo est soit :
- b/b, D+/D+, V+/V+
- b/b, D+/D, V+/V
- b/b, D+/D, V+/V+
Il est difficile de savoir car ces trois combinaisons donnent pratiquement la même tonalité de violet.
GRIS DOMINANT (deux allèles, G+ et G) - influe sur la structure des plumes.
Il y a donc trois possiblités pour les deux sexes :- G+/G+ : "type sauvage"
- G+/G : les plumes perdent toute leur capacité à diffracter la lumière (le bleu). Le vert devient kaki, on parle de "gris-vert", et le bleu devient gris.
- G/G :
Ce caractère de perte de la diffraction, et donc de la couleur bleue, est dominant.

ANTHRACITE (deux allèles connus, An+ et An) - influe sur la structure des plumes (en quantité)
Ce gène fonctionne de la même façon que le gène foncé, et a à peu près les mêmes effets. Il y a trois possibilités :- An+/An+ : "type sauvage", le corps reste clair
- An+/An : couleur légèrement foncée, un peu plus que vert foncé ou cobalt
- An/An : vert olive foncé en série verte, bleu très foncé à reflets mauves en série bleue. C'est surtout ce bleu qui permet de faire la différence avec les effets du gène foncé.
C'est un cas de codominance. L'allèle An est extrèmement rare.

(Photo copyright : Didier Mervilde)
BRONZE FALLOW (deux allèles connus fg+ et fg) - influe sur la syntèse de la mélanine
La chaine de synthèse de la mélanine est longue,avec de nombreux gènes, ce qui laisse la place à de nombreuses possibilités d'altération. Nous allons voir trois gènes différents, les "fallow", qui ont été identifiés chacun par un allèle, qui, porté en double exemplaire, empêchent la mélanine de devenir noire. Elle est donc brune, comme dans la couleur cinnamon, mais en encore plus clair. Ces trois gènes sont portés par des chromosomes non sexuels.- fg+/fg+ : type sauvage, mélanine noire
- fg+/fg : type sauvage, mélanine noire
- fg/fg : mélanine brune, yeux rouges sombre
Bronze fallow est une couleur récessive.
L'examen des plumes a révélé que la mélanine forme des granules plus petites et plus nombreuses que dans la couleur "cinnamon". Elle se répartit en outre en petits flocons.
Le fallow peut s'additionner avec les dilutions (en particulier l'allèle "dild" en double exemplaire), et "opaline" (allèle "op") pour donner des couleurs très proches des "ailes en dentelle" (voir Ino et cinnamon).
Une couleur très rare, le "Ino recessif", est considéré par certains auteurs comme dû à un allèle du gène "bronze fallow". En double exemplaire, cet allèle donnerait une couleur "Ino", entièrement dépourvue de mélanine. Mais cet allèle est extrèmement rare.
FALLOW ANGLAIS(deux allèles connus fe+ et fe)- influe sur la syntèse de la mélanine
Un autre gène porté par un chromosome non sexuel dont un allèle altère la nature de la mélanine quand il est en double exemplaire :- fe+/fe+ : type sauvage, mélanine noire
- fe+/fe : type sauvage, mélanine noire
- fe/fe : mélanine brune, yeux rouges clairs.
Ce caractère est donc récessif. Le fallow peut s'additionner avec les dilutions (en particulier l'allèle "dild" en double exemplaire), et "opaline" (allèle "op") pour donner des couleurs très proches des "ailes en dentelle" (voir Ino et cinnamon).

Fallow (Photo copyright : Brigitte Bodson)
FALLOW ECOSSAIS(deux allèles connus fs+ et fs)- influe sur la syntèse de la mélanine
- fs+/fs+ : type sauvage, mélanine noire- fs+/fs : type sauvage, mélanine noire
- fs/fs : mélanine brune, yeux rouges clairs avec un iris rose
Ce fallow est lui aussi une couleur récessive. Le fallow peut s'additionner avec les dilutions (en particulier l'allèle "dild" en double exemplaire), et "opaline" (allèle "op") pour donner des couleurs très proches des "ailes en dentelle" (voir Ino et cinnamon).
CORPS CLAIR DOMINANT (deux allèles connus, Cl+ et Cl) - influe la distribution de la mélanine
Voici un gène pour lequel on connaît deux allèles dont un est probablement le fruit d'une mutation survenue en élevage.Trois combinaisons possibles :
- Cl+/Cl+ : type sauvage. La mélanine est distribuée sur tout le corps
- Cl+/Cl : le ventre et le dos sont dépourvus de mélanine, les marques noires des ailes sont plus foncées (comme pour le "Texas clearbody", voir "Ino". Ce caractère est, comme son l'indique, dominant puisqu'il suffit d'un seul exemplaire de l'allèle pour qu'il soit exprimé).
- Cl+/Cl : le ventre et le dos sont dépourvus de mélanine, les marques noires des ailes sont plus foncées.
PIE DANOIS (deux allèles connus, r+ et r) - influe la distribution de la mélanine et du pigment jaune
- r+/r+ : type sauvage, la mélanine est présente sur tout le corps- r+/r : type sauvage, la mélanine est présente sur tout le corps
- r/r : des plumes sont totalement dépourvues de mélanine, selon un dessin alétoire (voir photo). L'oeil est noir, dépourvu d'iris, et la cire du bec rose chez les màles. Les ailes des oiseaux "verts" paraissent plus jaunes que chez les inos, ce qui indique que l'allèle influe aussi sur la distribution du pigment jaune...
La couleur pie danois est récessive.

PIE AUSTRALIEN (deux allèles connus, Pa+ et Pa) - influe la distribution de la mélanine
- Pa+/Pa+ : type sauvage, la mélanine est présente sur tout le corps- Pa+/Pa : des plumes sont totalement dépourvues de mélanine, selon un dessin plus ou moins semblable à celui de la photo ci-dessous. Les pattes sont souvent roses
- Pa/Pa : des plumes sont totalement dépourvues de mélanine, selon un dessin plus ou moins semblable à celui de la photo ci-dessous.Les pattes sont souvent roses
La couleur pie australien est dominante, mais de façon incomplète : l'importance de la surface décolorée varie selon les individus, et il n'est pas possible, juste en regardant la proportion de plumes décolorées, de savoir si ils ont l'allèle Pa en simple ou double exemplaire.
La proportion de plumes décolorées est probablement déterminée par un ou plusieurs autres gènes, qui n'ont pas encore été identifiés, bien que l'on soupçonne que certains soient liés au sexe.

PIE HOLLANDAIS (deux allèles connus, Pc+ et Pc) - influe la distribution de la mélanine
- Pc+/Pc+ : type sauvage, la mélanine est présente sur tout le corps- Pc+/Pc : des plumes sont totalement dépourvues de mélanine, selon un dessin plus ou moins semblable à celui du pie australien, à la différence que la zone claire du poitrail est plus haute, souvent jusqu'au masque. Les pattes sont souvent grises
- Pc/Pc : des plumes sont totalement dépourvues de mélanine, selon un dessin plus ou moins semblable à celui du pie australien, à la différence que la zone claire du poitrail est plus haute, souvent jusqu'au masque.Les pattes sont souvent grises
La couleur pie hollandais est dominante, mais, comme le pie australien, de façon incomplète : l'importance de la surface décolorée varie selon les individus, et il n'est pas possible, juste en regardant la proportion de plumes décolorées, de savoir si ils ont l'allèle Pc en simple ou double exemplaire.
La proportion de plumes décolorées est probablement déterminée par un ou plusieurs autres gènes, qui n'ont pas encore été identifiés, bien que l'on soupçone que certains soient liés au sexe.
Les oiseaux qui sont à la fois pie danois et pie hollandais (r/r Pc+/Pc ou r/r Pc/Pc) n'ont plus aucune trace de mélanine sur le plumage. Leurs yeux, par contre, sont entièrement noirs (un peu rougeàtres sous certaines lumières), sans iris blanc, ce qui permet de les distinguer des inos.
PERLéE (Gène Sp, mais on ne sait pas combien d'allèle sont en cause, ni même si un seul gène est impliqué) - influe la distribution de la mélanine sur le corps
Voici un gène pour lequel on connaît au moins deux allèles dont un est probablement le fruit d'une mutation survenue en élevage.Dans le cas le plus classique, on a trois combinaisons possibles :
- Sp+/Sp+ : type sauvage. La mélanine est distribuée sur tout le corps
- Sp+/Sp : Les ailes portent des taches claires en forme de perles. Ce caractère est codominant puisqu'il suffit d'un seul exemplaire de l'allèle pour provoquer la couleur perlée, mais qu'avec deux exemplaires... (voir ci-dessous)
- Sp/Sp : Le corps est entièrement dépourvu de mélanine, sauf les yeux qui restent noirs avec un iris blanc.
Selon certaines études([url=http://www.budgerigars.co.uk/specialist/spang/pilksp.html]ici par exmple">), les croisements de perlés ne donne pas les résultats attendus avec un schéma simple d'un gène à deux allèles. Il resterait donc encore des choses à découvrir sur la génétique de cette jolie couleur!
FACE NOIRE (deux allèles connus, bf+ et bf) - influe la distribution de la mélanine
Trois combinaisons possibles :- bf+/bf+ : type sauvage, la répartition de la mélanine est normale
- bf+/bf : type sauvage, la répartition de la mélanine est normale
- bf/bf : les ondulations noires passent du cou à la tête, en continant jusqu'au bec. Les spots se répètent aussi en remontant vers le bec. La tête semble foncée, mais pas noire. Il y a aussi de fines ondulations sur le ventre.
Ce caractère est récessif.

(Photo copyright : Didier Mervilde)
AILES SOMBRES (deux allèles connus, Dwi+ et Dwi) - influe sur la synthèse de la mélanine (en quantité)
Ce gène a été récemment découvert, et toute son hérédité n'est peut-être pas encore tout à fait bien connue. L'allèle Dwi ne peut s'exprimer que chez les oiseaux qui ont déjà le dessin des ailes "diluées" (par les allèles dild et dilgw en particulier). Il rstaure une partie de la dilution, en fonçant les ailes.- Dwi+/Dwi+ : les ailes restent claires
- Dwi+/Dwi : les ondulations sont un peu restaurées
- Dwi/Dwi : les ondulations sont davantage restaurées.
C'est un cas de codominance.
COULEURS RARES
Il existe encore d'autres couleurs, mais qui sont très rares, ou éteintes. On ne connaît pas forcément encore bien leur génétique. Parmi elles on peut citer :- Gris récessif, qui joue sur la structure des plumes comme gris dominant mais ne se transmet pas de la même manière
- Fallow australien, qui rend la mélanine brune
- Faded qui éclaircit joue sur la répartition de la mélanine, la diminuant surtout vers la tête et le haut du corps (qui deviennent vert ou bleu plus clair, avec un dégradé vers les plumes "normales"). voir ici

(Photo copyright : Didier Mervilde)
- Saddlebacked : qui éclaircit la mélanine au niveau des épaules et du dos (les ondulations noires deviennent grises)
- Mottled : des plumes sont dépourvues de mélanine, un peu plus à chaque mue.
- Frosted : les plumes de la poitrine ne portent plus de mélanine, ce qui donne aux bleux un aspects "givré"
- Ailes brunes/sepia : ondulations des ailes d'un brun terne. Récessif. Semble disparu depuis la seconde guerre mondiale
(voir ici)
- Misty : la couleur du corps est diluée légèrement. Caractère codominant (ceux qui ont deux exemplaires de l'allèle récemment découvert sont éclaircis d'environ 25%) (voir ici)

(Photo copyright : Didier Mervilde)
Une partie de ces couleurs rares sont visibles ici.
Sources :
Wikipedia
Le site de Didier Mervilde
Mutavi research group
Forum Calopsittes et Psittacidés
merci à Didier Mervilde, Takara, Brigitte Bodson et Pascale Maitre de m'avoir autorisée à utiliser leurs photos!
Lexique de génétique pour ceux qui veulent apprendre des termes supplémentaires :
- homozygote : les deux allèles que portent l'individu sont identiques. C'est le cas des "simple facteur"
- hétérozygote : les deux allèles que portent l'individu sont différents. C'est le cas des "double facteur", mais aussi de ceux qui portent deux allèles de type sauvage.
- crossing-over : "échange de morceaux" entre chromosomes au moment de la fabrication des gamètes.
- génotype : ensemble des allèles portés par l'individu. On écrit uniquement ceux des gènes qui nous intéressent, c'est à dire les gènes pour lesquels on sait que l'oiseau porte au moins un allèle différent du "type sauvage".
- phénotype : aspect extérieur de l'oiseau, couleur et forme de ses plumes, forme de son bec etc
- autosome : chromosome non sexuel
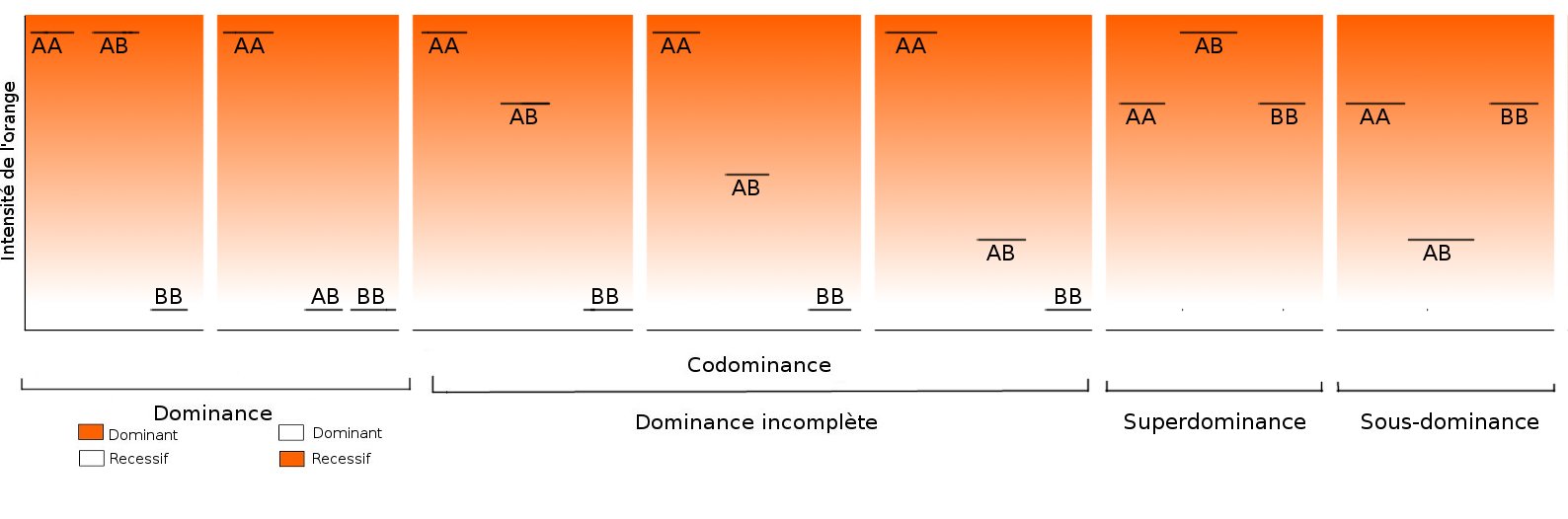
Récapitulatif de toutes les relations possibles entre allèles, avec un exemple imaginaire de synthèse de pigment orange, et un gène à deux allèles : A et B.
AA et BB sont homozygotes
AB est hétérozygote.
Entre Zcin et Zino.............3%
Entre Zcin et Zop............33%
Entre Zop et Zino............30%
Entre Zino et Zsl...............7%
Entre Zop et Zsl..............40%
Entre Zino et Zsl..............10%
Entre Bleu et Foncé..........14%[/i]